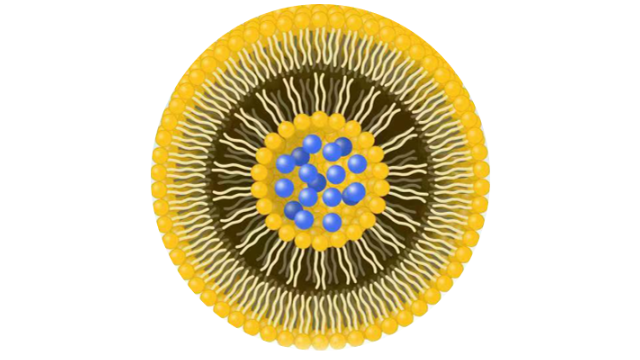
因此,可以实现靶向和长 循环的双重好处。 免疫脂质体是利用抗体或其片段与脂质体之间的各种类型的连锁来制备的。根据制备方法的不同, 可以在脂质上进行连接, 然 后脂质可用于制造脂质体或可以在脂质体上进行连接。 常用的键合类型是抗体和脂质体之间的共价和非共价偶联。在共价偶联中, 氨基(酰胺键形成)或巯基(马来酰亚胺反应) 是偶联过程的主要活性位点。然而, 在非共价偶联中, 用生物素修饰的脂质体制备脂质体, 靶向蛋白分子附着在脂质体上。增加循环半衰期, 靶向特异性和**小化药物损失和降解是免疫脂质体的主要优点。 除了有前景 的应用之外, 免疫脂质体还有一个主要缺点, 即由于反复注射, 可以观察到免疫原性和循环***率的增加。小于80纳米的免疫脂质体(作为有效递送的要求)可能会从肿瘤部位迅速消除。相变温度对脂质体的影响。长沙脂质体载药惰性气体
microRNA脂质体
microRNA是真核细胞中发现的短(约22mer)非编码RNA,通过结合互补的mRNA序列发挥生物调节剂的作用。miRNA以初级miRNA的形式从其编码的核基因转录,其长度为数百个核苷酸。RNaseIII酶,Drosha,将初级miRNA加工成pre-miRNA(长度为70个核苷酸),携带一个特征的发夹环。然后pre-miRNA移动到细胞质中,在那里RNaseIII酶Dicer产生成熟的miRNA和乘客链。***,成熟的miRNA被整合到RNAi诱导的沉默复合体中,以降解它们的靶mRNA。由DOTMA、胆固醇和vitaminETPGS1k琥珀酸盐组成的阳离子脂质体被证明可以有效递送pre-miRNA-133b,导致A549非小肺*细胞中成熟miRNA-133b的表达比对照组细胞增加2.3倍,Mcl-1蛋白的表达减少1.8倍。经尾静脉注射含有pre-miRNA-133b的阳离子脂质体(1.5mg/kg)的ICR小鼠肺组织中成熟miRNA-133b的表达比接受含有紊乱的pre-mirna的阳离子脂质体的小鼠高52倍。 河北脂质体载药试剂PEG2000是一种聚乙二醇(PEG)衍生物,常用于脂质体的表面修饰。
脂质体疫苗通常在已知疫苗中使用纯化抗原或减毒病原体作为免疫原。然而,长期的免疫反应可能不会由纯化抗原诱导,甚至有时根本不会诱导反应。另一方面,减毒疫苗可以在免疫的患者中产生应答。然而,递送包裹在脂质体内的抗原可诱导长期应答,这在某些抗原的直接免疫中没有观察到。研究表明,恶性细胞的细胞膜可以形成包封潜在抗原的脂质体。文献报道了包封在脂质体中的肽作为**疫苗的***应用能力。有研究评估了BLP25(一个含有合成人MUC1肽的25个氨基酸序列)作为**疫苗的能力。用二硬脂酰磷脂酰胆碱、胆固醇和二肉豆醇酰磷脂酰甘油(摩尔比3:1:25)中含有的单磷脂酰脂A(1%w/w)制备脂质体,然后与脂偶联和非偶联的MUC1肽结合。C57BL/6小鼠免疫分别采用肽相关脂质体、肽与无肽脂质体混合、脂肽单独免疫。结果表明,脂质体制剂对免疫应答有深远的影响。与物理相关的脂质体观察到强烈的免疫反应(抗原特异性t细胞细胞反应),而与肽混合的无肽脂质体或单独的脂肽则没有。体液免疫反应受到关联性质的***影响,这可以通过表面暴露的肽脂质体诱导muc1特异性抗体来证明。因此可以通过调整脂质体药物传递系统来诱导优先细胞反应这提出了一个假设即不同的脂质体配方刺激不同的免疫途径。
脂质体靶向递送中甘露糖配体修饰由于在巨噬细胞上发现了甘露糖受体,因此甘露糖已被用于修饰阳离子脂质体以供巨噬细胞递送。为了抑制由活化的巨噬细胞诱导的破骨细胞生成,将甘露糖基化阳离子脂质体与双链寡核苷酸NFkB诱饵络合。甘露糖阳离子脂质体/NFkB诱饵复合物有效诱导NFkB活化并抑制肿瘤坏死因子-a的产生。在另一项研究中,巨噬细胞靶向NFkB诱饵装载在甘露糖基化阳离子脂质体中,用于预防脂多糖诱导的肺部炎症。气管内给药后,甘露糖标记的阳离子脂质体/NFkB诱饵复合物***下调NFkB的表达,减少肿瘤坏死因子-a和白细胞介素-1b的释放。研究人员研究了茴香酰胺修饰的阳离子脂质体将寡核苷酸靶向递送至表达sigma受体的细胞的能力。剪接开关寡核苷酸(SSOs)是一种单链寡核苷酸,可与剪接位点或剪接增强子结合,阻断内源性剪接机制的通路,并产生成熟mRNA的替代版本。在肺转移小鼠模型中,全身给药装载Bcl-xSSO的茴香胺修饰阳离子脂质体可降低**生长。增强成像性能,荧光标记的定量分析,探索药物的药代动力学以及研究药物的靶向性等。

对筛选的阳离子脂质进行了研究,以6.25mg/kg的剂量给食蟹猴全身给药载脂蛋白B特异性siRNA,据报道,在2周内,肝组织中载脂蛋白B的表达减少了50%以上。近年来,研究人员合成了多种阳离子脂质体,并试图找到一种可有效递送质粒DNA的阳离子脂质体组合物。在新合成的阳离子中,N',N',-dioctadecyl-N-4,8-diaza-10-aminodecanoylglycine(DODAG)制成的阳离子纳米脂质体对质粒DNA的转染效率比较高。此外,DODAG比转染试剂Lipofectamine2000更有效地将质粒DNA传递到OVCAR-3和HeLa细胞系。相比之下,基于理性的新型阳离子脂质预测是基于这样一种假设,即阳离子脂质在内吞作用后可以与内体膜的天然阴离子脂质相互作用,锥形脂质会诱导双层膜的破坏。为了设计能够提高转染效率的阳离子脂质,作者控制了脂质头基团、碳氢化合物结构域和连接体。由于AS-ODNs可以下调某些RNA并抑制靶蛋白的表达,因此它们被认为具有作为核酸药物的潜力。长沙脂质体载药惰性气体
脂质体的缓释作用可以减少给药频率。长沙脂质体载药惰性气体
脂质体中辅助脂质中性脂也经常被用作阳离子脂质体的助手。例如,已知中性脂质1,2-二油基-asn-甘油-3-磷酸乙醇胺(DOPE)在胞吞作用后参与内体逃逸,胆固醇(一种内源性脂质)可以插入脂质双层之间以增加纳米颗粒的刚性。为了增加体内稳定性,一种非常普遍的方法包括插入聚乙二醇(PEG)偶联的中性脂质,对纳米颗粒进行聚乙二醇化。此外,中性辅助性脂质,如DOPE已被用于提高阳离子脂质体的递送效率。DOPE提高核酸递送效率的生物物理机制仍在研究中。**近的一项研究报道,含有DOPE的脂质单层呈现不规则的豆状结构域,而缺乏DOPE的脂质单层呈现均匀的表面。除DOPE外,其他中性脂质,包括N-十二烷酰基肌氨酸,已被报道可提高阳离子脂质体的基因递送效率。长沙脂质体载药惰性气体