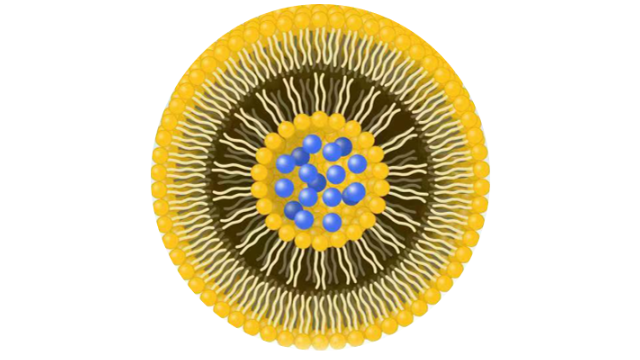
siRNA脂质体
RNA干扰(RNAi)途径允许siRNA和miRNAs负向调节蛋白表达。siRNA是21~23对核苷酸组成的双链RNA,可诱导同源靶mRNA沉默。为了发挥作用,双链siRNA分裂成两个单链RNA:乘客链和引导链。乘客链被argonaute-2蛋白降解,而引导链则被纳入RNAi诱导的沉默复合体中,该复合体结合与引导链互补的mRNA并将其切割。siRNA似乎具有***多种疾病的巨大潜力,因为它们可以很容易地下调各种靶mRNA,而不考虑它们的位置(即在细胞核或细胞质中),并且它们的特异性结合表明它们比传统化学药物诱导的副作用更少。作为一种新型的基于核酸的***策略,siRNA***与传统的化学药物相比具有许多优势。然而,为了促进基于siRNA的***方法的发展,必须克服一些挑战,包括需要识别适当的靶基因和开发优化的递送系统。许多研究人员试图利用阳离子脂质体提高siRNA的细胞递送和基因沉默效率。例如,由DC-6-14、DOPE和胆固醇组成的阳离子脂质体被用于递送萤火虫荧光素酶特异性的siRNA。当阳离子脂质体与siRNA持续剧烈搅拌混合时,转染效率提高,说明将siRNA加载到阳离子脂质体上的方法可以调节转染效率。siRNA脂丛的***应用因靶蛋白而异。 增强成像性能,荧光标记的定量分析,探索药物的药代动力学以及研究药物的靶向性等。脑靶向脂质体载药技术服务公司

microRNA脂质体
microRNA是真核细胞中发现的短(约22mer)非编码RNA,通过结合互补的mRNA序列发挥生物调节剂的作用。miRNA以初级miRNA的形式从其编码的核基因转录,其长度为数百个核苷酸。RNaseIII酶,Drosha,将初级miRNA加工成pre-miRNA(长度为70个核苷酸),携带一个特征的发夹环。然后pre-miRNA移动到细胞质中,在那里RNaseIII酶Dicer产生成熟的miRNA和乘客链。***,成熟的miRNA被整合到RNAi诱导的沉默复合体中,以降解它们的靶mRNA。由DOTMA、胆固醇和vitaminETPGS1k琥珀酸盐组成的阳离子脂质体被证明可以有效递送pre-miRNA-133b,导致A549非小肺*细胞中成熟miRNA-133b的表达比对照组细胞增加2.3倍,Mcl-1蛋白的表达减少1.8倍。经尾静脉注射含有pre-miRNA-133b的阳离子脂质体(1.5mg/kg)的ICR小鼠肺组织中成熟miRNA-133b的表达比接受含有紊乱的pre-mirna的阳离子脂质体的小鼠高52倍。 广东昆明脂质体载药脂质体是由多种组分构成的,主要包括:磷脂质、胆固醇、表面活性剂和PEG2000等。

脂质体靶向递送中RGD配体修饰尽管阳离子脂质体具有在体内递送核酸的潜力,但其递送到特定靶点仍然是一个主要挑战。为了增强携带核酸的阳离子脂质体在靶组织中的分布,研究人员用多肽和小分子修饰了脂质体表面。例如,研究了Arg-Gly-Asp(RGD)肽修饰的脂质体增强核酸向整合素受体表达细胞传递的能力。负载P糖蛋白特异性siRNA的RGD修饰阳离子脂质体对整合素受体表达的人乳腺*MCF7/A细胞的递送率更高,导致P糖蛋白的***沉默。与此一致的是,分子成像显示,与小鼠模型的邻近正常组织相比,MCF7/A**组织中RGD修饰的阳离子脂质体和siRNA的分布更高。在**近的一项研究中,用环RGD和辛精氨酸修饰脂质体表面,以利用环RGD的整合素受体结合效应和辛精氨酸的细胞穿透效应。双配体修饰的阳离子脂质体增加了整合素avb3表达细胞的细胞摄取,并且更有效地转染荧光素酶编码质粒DNA。
脂质体共价连接药物-脂质偶联载***式通过连接剂将药物分⼦与脂质共价连接是另⼀种在脂质体内装载药物的有效策略,例如Mepact。MDP是主要⾰兰⽒阳性菌细胞壁的组成部分,具有****应答的作⽤。由于MDP是⽔溶性低分⼦量分⼦,其脂质体在储存过程中存在包封效率低和药物泄漏等问题。为了提⾼MDP的脂溶性,通过肽间隔剂将MDP与PE连接,合成MTP-PE(muramyltripeptide-phosphatidylethanolamine)。在⽤⽣理盐⽔重建冻⼲产物(MTP-PE,POPC和OOPS)时,MTP-PE的两亲分⼦嵌⼊脂质体的膜双层。脂质体内存在MTP-PE,未发现游离MTP-PE。Vyxeos采⽤被动加载和主动加载相结合的⽅法,这是⾸个被批准在同⼀囊泡中加载两种不同药物(阿糖胞苷和柔红霉素)的脂质体。简⽽⾔之,当脂质泡沫与Cu(葡糖酸盐)2、三⼄醇胺(TEA)、pH7.4和阿糖胞苷溶液⽔合时,阿糖胞苷被被动地封装到脂质体中。经过减浆和缓冲液交换以去除未包封的药物和Cu(葡糖酸盐)2/TEA后,中性pH的柔红霉素缓冲液与载糖胞苷脂质体孵育。一些常用于标记脂质体的荧光染料包括:DiO、DiI、Rhodamine PE、NBD、BODIPY、Cy3和Cy5等。
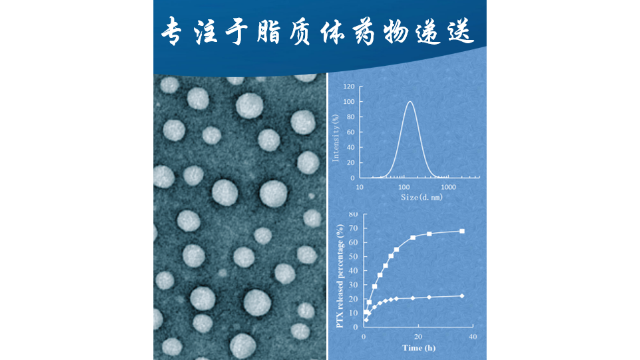
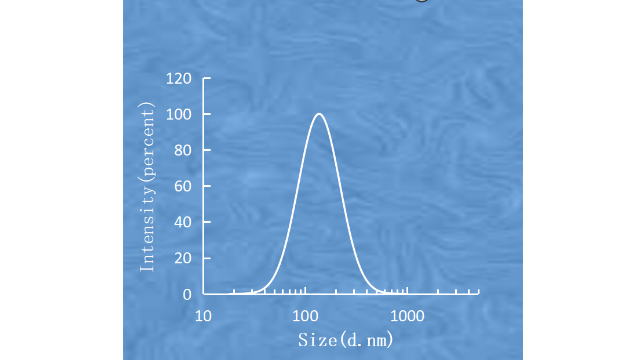
酸性环境(pH值2.0-4.0)通常⽤于产⽣⽤于活***物装载的跨膜pH梯度。在37℃和pH2.0条件下,SM/Chol脂质体(55/45,mol/mol)的⽔解速率⽐DSPC/Chol脂质体慢约100倍。此外,含有SM/Chol的脂质体表现出比较好的药代动⼒学特性,即增加循环时间并增强药物向靶组织的递送。胆固醇(Chol)是脂质体双分⼦层的另⼀个主要成分,⼏乎可以⽤于所有的商业产品。Chol的加⼊可以促进脂链的堆积和双分⼦层的形成,调节膜的流动性/刚性,并进⼀步影响药物释放、脂质体的稳定性和胞外分泌动⼒学。对于Shingrix(带状疱疹疫苗,含有糖蛋⽩E抗原和AS01B脂质体佐剂系统)的产物,Chol可以避免QS21(AS01B佐剂系统中的免疫增强剂之⼀)以2:1的⽐例(Chol:QS21,w/w)⽔解。对于AmBisome的产物,与⾮甾醇相⽐,Chol降低了脂质体制剂的毒性。Chol对双分⼦层性质的影响是浓度依赖性的。据报道,低浓度(2.5mol%)和⾼浓度(>30mol%)的Chol对脂质双分⼦层的性质影响不⼤。5<Cholmol%<30的Chol的“冷凝效应”或“有序效应”导致颗粒⼤⼩从220nm逐渐增⼤到472nm,膜的流动性降低,药物释放减少。除了Chol,其他与Chol结构相似的甾醇,如⻩体酮、⻨⻆甾醇和⽺⽑甾醇,也被研究⽤于调节膜的刚性和稳定性。被动载药⽅法是在脂质体制备过程中对药物进⾏包封的方法。脑靶向脂质体载药技术服务公司
将荧光标记引入载药脂质体的作用有荧光标记的定位和跟踪,药物释放的实时监测。脑靶向脂质体载药技术服务公司
脂质体各组分对核酸递送效率的影响对于使用阳离子脂质体开发核酸***剂,一个先决条件是必须将核酸适当地递送到靶细胞并到达适当的亚细胞区室(例如,细胞质或细胞核)。已知阳离子脂质体的递送效率会受到阳离子脂质和辅助脂质类型及其组成的影响。阳离子脂质是纳米粒子的**成分,具有一个带正电的头基和一个或两个由碳氢链或类固醇结构组成的疏水尾区的共同结构。Felgner和同事报道了N-[1-(2,3-二聚氧基)丙基]-N,N,N-三甲基氯化铵(DOTAP)的合成,其具有一个单价阳离子头和两个碳氢化合物尾部,并用于制备小的单层脂质体。他们将DNA包裹的脂质体转染到小鼠L细胞中,并证明阳离子脂质中和了带负电荷的DNA,使阳离子脂质体有更好的机会与带负电荷的细胞膜相互作用。从那时起,各种阳离子脂质和基于脂质的纳米颗粒被设计和评估用于核酸的细胞递送,包括DNA,siRNA,miRNA和AS-ODN。这些新的阳离子脂质已经通过文库技术和基于理性的预测相结合的方法被鉴定出来。对类脂类材料文库的筛选产生了由十个碳和两个烷基链组成的阳离子脂质,发现其比其他候选物更有效。
脑靶向脂质体载药技术服务公司